環(huán)境工程投稿論文格式參考:畜禽糞污厭氧消化產(chǎn)甲烷效能的關(guān)鍵影響因素研究進(jìn)展
關(guān)鍵詞:畜禽糞污;厭氧消化;產(chǎn)甲烷古菌;產(chǎn)甲烷潛力;導(dǎo)電材料
作者:孫廣東;馬俊怡;郝 書(shū);董 賀;趙心茹;柴強(qiáng)龍;黨 巖
作者單位:北京林業(yè)大學(xué);
摘 要:在畜禽養(yǎng)殖方式逐漸規(guī)模化的背景下,規(guī)模化養(yǎng)殖所產(chǎn)生的糞污具有集中排放量大、處置需求強(qiáng)的特點(diǎn),這為 利用厭氧消化技術(shù)處置畜禽糞污來(lái)實(shí)現(xiàn)沼氣產(chǎn)量、產(chǎn)能效益最大化提供了契機(jī),完全契合我國(guó)能源結(jié)構(gòu)優(yōu)化、碳中和 等戰(zhàn)略的需要。然而,由于畜禽糞污組分復(fù)雜、厭氧消化調(diào)控操作條件繁多以及技術(shù)自身反應(yīng)速率限制等原因影響, 厭氧消化技術(shù)在處置畜禽糞污過(guò)程中存在產(chǎn)氣效率低、過(guò)程穩(wěn)定性差的缺陷,掌握其產(chǎn)甲烷效能關(guān)鍵影響因素變得尤 為重要。該文分析介紹了不同畜禽糞污的特性以及影響其厭氧消化產(chǎn)甲烷潛力的原因,綜述了近年來(lái)畜禽糞污厭氧 消化的溫度、pH、氨氮等主要環(huán)境因子以及進(jìn)料負(fù)荷、進(jìn)料方式等外部調(diào)控因素對(duì)產(chǎn)甲烷效能、微生物群落豐度的影 響研究,探討了投加功能菌劑、導(dǎo)電材料等輔助方式對(duì)系統(tǒng)產(chǎn)甲烷效能提升的作用以及系統(tǒng)微生物的反饋機(jī)制,對(duì)后 續(xù)的重點(diǎn)研究方向進(jìn)行了展望,以期為未來(lái)的畜禽糞污厭氧消化技術(shù)理論研究與工程應(yīng)用提供參考。
0 引 言
由于我國(guó)居民飲食結(jié)構(gòu)中畜禽產(chǎn)品占比逐漸增 大,畜禽養(yǎng)殖市場(chǎng)呈現(xiàn)出穩(wěn)定增長(zhǎng)的趨勢(shì),畜禽糞污 排放量與日俱增。根據(jù)農(nóng)業(yè)農(nóng)村部等相關(guān)數(shù)據(jù)統(tǒng) 計(jì),全國(guó)每年產(chǎn)生畜禽糞污總量近 40 億 t,約為生活 垃圾年產(chǎn)量的 16 倍[1]。畜禽糞污中有機(jī)質(zhì)含量為 30%~70%,有效處理率不到 50%,畜禽養(yǎng)殖業(yè)水污染 物排放量的化學(xué)需氧量(COD)約為 1000. 53 萬(wàn) t,占 全國(guó)水污染物總排放量的 46. 67%[2] 。畜禽糞污是具 有巨大應(yīng)用潛力的碳源,其等效二氧化碳排放量約 為 1. 23 億 t,為生活垃圾處理碳排放量的 2. 5 倍。厭 氧消化技術(shù)通過(guò)微生物的代謝活動(dòng)將廢棄有機(jī)物穩(wěn) 定化,同時(shí)伴有甲烷和二氧化碳產(chǎn)生,具有處置畜禽 糞污減量無(wú)害化、產(chǎn)生沼氣可再生能源的雙重優(yōu)勢(shì), 引起各國(guó)關(guān)注[3, 4] 。隨著我國(guó)養(yǎng)殖方式從分散飼養(yǎng)向 集約化和現(xiàn)代化的方式快速轉(zhuǎn)變,所產(chǎn)畜禽糞污集 中化程度不斷提高,這為厭氧消化技術(shù)推廣后的設(shè) 施規(guī)模化、沼氣能源產(chǎn)量化、產(chǎn)能效益最大化提供了 可能。
然而,畜禽糞污厭氧消化產(chǎn)甲烷過(guò)程歷經(jīng)水解、 酸化、乙酸化和甲烷化等階段,是一種由具有不同代 謝機(jī)制的微生物共同參與的生物化學(xué)反應(yīng)過(guò)程,系 統(tǒng)中微生物之間的協(xié)同合作既微妙又復(fù)雜[5] 。厭氧 消化過(guò)程涉及微生物主要包括水解酸化細(xì)菌和產(chǎn)甲 烷 古 菌 ,至 少 涵 蓋 了 20 個(gè) 門(mén) 的 細(xì) 菌 和 5 個(gè) 目 的 古 菌[6, 7] 。產(chǎn)甲烷古菌作為厭氧消化過(guò)程關(guān)鍵微生物, 是嚴(yán)格的專(zhuān)性厭氧菌,只能利用有限的簡(jiǎn)單有機(jī)物, 主要為乙酸、氫氣和少數(shù)簡(jiǎn)單有機(jī)物[8] 。產(chǎn)乙酸菌和 產(chǎn)甲烷古菌之間互營(yíng)關(guān)系成為影響厭氧消化最重要 的因素,這兩類(lèi)微生物的不平衡是導(dǎo)致厭氧消化系 統(tǒng)不穩(wěn)定的首要原因[9] 。傳統(tǒng)的厭氧消化存在產(chǎn)氣 效率低、過(guò)程穩(wěn)定性差的缺陷,常常遭遇超負(fù)荷、氨 抑制等誘發(fā)的過(guò)程失穩(wěn)現(xiàn)象。厭氧消化系統(tǒng)一旦失 穩(wěn),二次啟動(dòng)運(yùn)行通常需要較長(zhǎng)時(shí)間恢復(fù),從而為其 所服務(wù)的養(yǎng)殖企業(yè)或固廢環(huán)保企業(yè)帶來(lái)極大的環(huán) 保、經(jīng)濟(jì)風(fēng)險(xiǎn)。因此,探明影響畜禽糞污厭氧消化過(guò) 程穩(wěn)定以及促進(jìn)消化產(chǎn)甲烷效率的關(guān)鍵因素,對(duì)厭 氧消化技術(shù)可持續(xù)性發(fā)展具有重要意義。
在化石能源逐漸枯竭的背景下,依托厭氧消化 技術(shù)實(shí)現(xiàn)畜禽糞污的資源化高效利用,是提升碳回 收和生物質(zhì)能生產(chǎn)效率的重要舉措,對(duì)助力實(shí)現(xiàn)我 國(guó)的“雙碳目標(biāo)”意義重大[10] 。本文分析了以不同畜 禽糞污為底物的厭氧消化產(chǎn)甲烷效能與潛力,總結(jié) 了畜禽糞污厭氧消化產(chǎn)甲烷過(guò)程中不穩(wěn)定的關(guān)鍵環(huán) 境因素和干擾機(jī)制,探討了進(jìn)料負(fù)荷和不同進(jìn)料方 式對(duì)產(chǎn)甲烷效能的影響,對(duì)后期畜禽糞污厭氧消化 技術(shù)的重點(diǎn)研究方向進(jìn)行了展望,以期為該技術(shù)的 基礎(chǔ)研究和工程化應(yīng)用提供參考。
1 不同底物類(lèi)型厭氧消化產(chǎn)甲烷效能與潛力分析
牛糞、豬糞和雞糞是我國(guó)主要的 3 種畜禽糞污, 分 別 占 畜 禽 糞 污 總 產(chǎn) 量 的 45. 2%、39. 2% 和 15. 6%[11] 。由于畜禽消化特征的差異和飼料成分、飼 養(yǎng)方式以及糞污收集方式等外部因素,各畜禽糞污 的成分結(jié)構(gòu)、理化性質(zhì)、產(chǎn)甲烷潛能各不相同[12] 。各 畜禽糞便的成分對(duì)比見(jiàn)表 1。
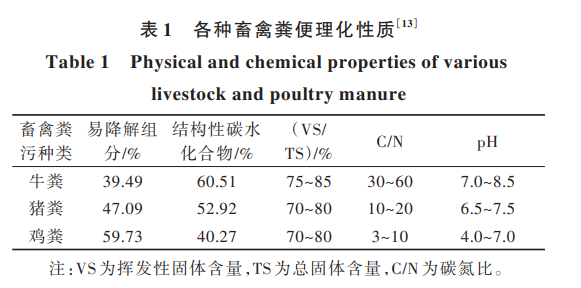
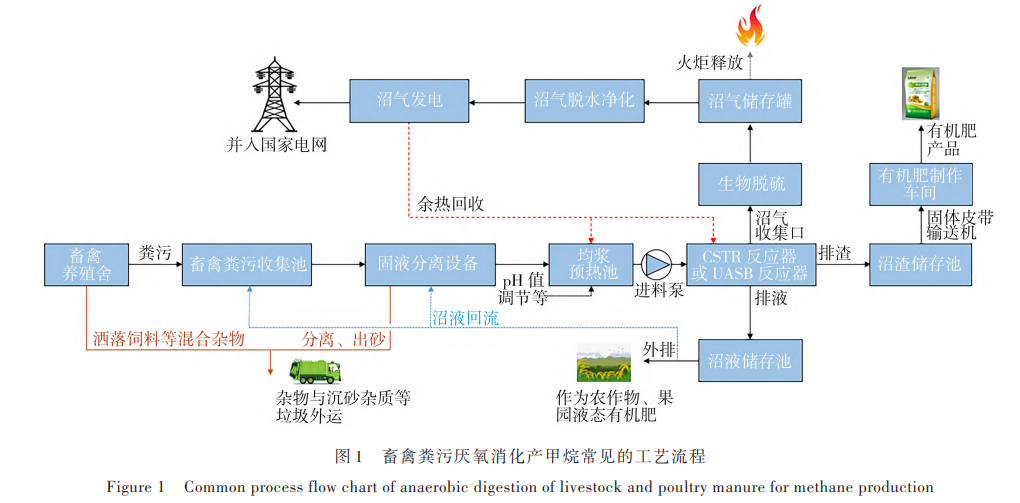
糞污 底 物 中 易 降 解 組 分 含 量 由 高 到 低 依 次 為 雞 糞 (59. 73%) 、豬 糞 (47. 08%) 和 牛 糞 (39. 49%)。 常 見(jiàn) 的 畜 禽 糞 污 厭 氧 消 化 產(chǎn) 甲 烷 工 藝 為 連 續(xù) 攪 拌 式 反 應(yīng) 器(CSTR)工 藝 、升 流 式 厭 氧 污 泥 床 反 應(yīng) 器(UASB)工 藝 ,具 體 工 藝 流 程 見(jiàn) 圖 1。
1. 1 牛糞污的厭氧消化產(chǎn)甲烷效能與潛力
牛類(lèi)反芻動(dòng)物以草飼料為主,其瘤胃是一個(gè)天 然發(fā)酵罐,瘤胃內(nèi)部微生物體系具有強(qiáng)大的木質(zhì)纖 維素水解酸化能力,將復(fù)雜的木質(zhì)纖維素生物質(zhì)轉(zhuǎn) 化為可溶性有機(jī)物和揮發(fā)性脂肪酸(VFAs),通常能 夠在 2 d 內(nèi)降解 60%以上的纖維[14]。由于牛飼料中 易降解部分已經(jīng)被牛瘤胃中微生物利用,牛糞中易 降解組分與典型的雞糞、豬糞相比含量最低,含有超 過(guò)總固體含量 50% 的不易降解有機(jī)組分,包括纖維 素、半纖維素和木質(zhì)素,其復(fù)雜的結(jié)構(gòu)而阻礙了牛糞 在厭氧消化中的水解,使其成為限速步驟。根據(jù)牛 糞化學(xué)成分分析結(jié)果,理論上牛糞的厭氧消化甲烷 生產(chǎn)潛力值為 206~219 mL/g TS,實(shí)際實(shí)驗(yàn)中產(chǎn)甲烷 潛 能 為 130~139 mL/g TS,僅 為 前 者 的 60%~ 70%[15, 16] 。
提高牛糞厭氧消化的效能,通常使用預(yù)處理、共 消化、添加劑等途徑來(lái)解決。通過(guò)預(yù)處理可以分解 復(fù)雜的有機(jī)物,使木質(zhì)纖維素化合物更容易被水解 從而提高厭氧消化效能;共消化是通過(guò)將不同性質(zhì) 的有機(jī)廢棄物混合,以提高厭氧消化效能。生物酶 添加劑能夠?qū)㈦y降解的物質(zhì),分解為更易被微生物 利用的小分子有機(jī)物,從而提升厭氧消化的速率。 Fernandez 等[17] 研究發(fā)現(xiàn)在對(duì)牛糞進(jìn)行機(jī)械破碎和混 合的預(yù)處理后,甲烷的產(chǎn)量可以提高 12%。與其他畜 禽糞污相比,牛糞中所含的氮化合物含量較低,但結(jié) 構(gòu)性碳水化合物含量最高,屬于富碳基質(zhì),其較高的 C/N(30~60)也是阻礙厭氧消化的主要障礙之一[18]。 當(dāng)牛糞廢水與其他有機(jī)組分進(jìn)行共消化時(shí),其強(qiáng)大 的緩沖能力更有利于提高甲烷產(chǎn)量,即牛糞的存在 會(huì)促進(jìn)給定底物的厭氧消化過(guò)程。Xing 等[19] 研究發(fā) 現(xiàn)當(dāng)牛糞廢水與餐廚垃圾以 1∶2. 5 的比例混合時(shí),甲 烷的產(chǎn)率最高可達(dá)到 2. 71 L(/ L·d)。在牛瘤胃內(nèi)部 的微生物含有產(chǎn)甲烷古菌,其數(shù)量占瘤胃微生物總 量的 2%~4%。目前已有研究分離出的瘤胃液中產(chǎn)甲 烷 古 菌 主 要 有 4 個(gè) 屬 :甲 烷 八 疊 球 菌 屬 Methanosarcina、甲 烷 微 菌 屬 Methanomicrobium、甲 烷 短 桿 菌 Methanobrevibacter 和 甲 烷 桿 菌 屬 Methanobacterium[20, 21]。當(dāng)瘤胃液與牛糞以 3∶7 的比 例混合時(shí),牛糞中的微生物與瘤胃液中的微生物各 自發(fā)揮優(yōu)勢(shì),分工協(xié)作,甲烷產(chǎn)率最大值達(dá)到 169 mL/ g VS [22] 。
1. 2 豬糞污的厭氧消化產(chǎn)甲烷效能與潛力
豬糞成分復(fù)雜,包括蛋白質(zhì)、脂肪、有機(jī)酸、纖維 素 、半 纖 維 素 等 ;含 氮 較 多 ,C/N 約 為 14;脂 肪 含 量 (11. 18%)顯著高于牛糞、雞糞[13] 。相對(duì)于保育豬、生 長(zhǎng)期豬的豬糞,妊娠母豬的豬糞含有更多的纖維素、 木質(zhì)素和芳香族化合物等難降解有機(jī)物,這些物質(zhì) 高度腐殖化,難生物降解,保育豬糞便、生長(zhǎng)期豬糞便和妊娠母豬糞便的產(chǎn)甲烷值分別為 273,272,230 mL/g VS,且保育豬糞便和生長(zhǎng)期豬糞便的產(chǎn)甲烷過(guò) 程十分相近[23] 。
由于養(yǎng)殖模式的原因,不少豬場(chǎng)使用水泡糞清 糞工藝清理豬糞便,使得每頭豬平均每天產(chǎn)生 4~8 L 廢水,主要包括洗滌、糞便以及尿液等成分;豬糞受 沖欄洗滌水的摻混,所形成豬糞廢水具有排放水量 大,且時(shí)間相對(duì)集中的特點(diǎn)[24, 25] 。在一定范圍內(nèi),隨 著豬糞廢水 TS 的增加,厭氧發(fā)酵產(chǎn)氣效能也增加;若 豬糞廢水 TS 含量較低時(shí),發(fā)酵系統(tǒng)內(nèi)酸濃度低,產(chǎn)甲 烷古菌群底物不足,雖然產(chǎn)氣速率快,但容積產(chǎn)氣效 率低;若豬糞廢水 TS 過(guò)高,形成的水泡糞流動(dòng)性差, 給豬舍的糞污管理帶來(lái)麻煩。選擇合適的水泡糞 TS 含量對(duì)其厭氧消化尤為重要。陳欣等[26] 研究豬糞廢 水 TS 在 2%、6%、10% 和 14% 4 個(gè)梯度下厭氧消化試 驗(yàn)下,發(fā)現(xiàn) TS 對(duì)豬糞厭氧消化過(guò)程及甲烷產(chǎn)率有較 大影響,TS 為 6% 和 10% 的甲烷產(chǎn)率較高,當(dāng) TS 進(jìn)一 步提高至 14% 時(shí),物料中過(guò)高 VFAs 以及氨氮濃度對(duì) 產(chǎn)甲烷古菌群活性產(chǎn)生抑制作用。
1. 3 雞糞污的厭氧消化產(chǎn)甲烷效能與潛力
雞禽類(lèi)常以谷物為飼料,飲水少,加之雞禽類(lèi)的 消化道僅為其體長(zhǎng)的 7 倍,比其他畜禽的消化道短, 飼料在體內(nèi)停留僅約 4 h,營(yíng)養(yǎng)物質(zhì)的消化吸收率很 低 ,40%~70% 營(yíng) 養(yǎng) 物 質(zhì) 會(huì) 隨 糞 便 排 出 體 ,肥 分 濃 厚[27, 28] 。理論上每克干雞糞全部降解可以產(chǎn)生 421~ 520 mL,實(shí)際實(shí)驗(yàn)中產(chǎn)甲烷潛能為 250~370 mL/g TS, 比典型的牛糞、豬糞產(chǎn)甲烷潛能高[29, 30] 。
雞糞屬于高氮物質(zhì),相對(duì)于其他畜禽糞便,含有 更多的蛋白質(zhì)、氨基酸及尿酸,其蛋白質(zhì)組分最高, 為牛糞、豬糞的 2 倍[13] 。在厭氧消化處理過(guò)程中,有 機(jī)氮轉(zhuǎn)化為氨氮(包括 NH+ 4 和游離氨 NH3 )的效率為 62. 6%~80. 3%,故高固體雞糞厭氧消化面臨著更大 的氨抑制風(fēng)險(xiǎn),因此常控制雞糞厭氧消化 TS 含量在 10% 以下[31] 。最初的雞糞厭氧消化處理研究主要在 低濃度(0. 5%~3%)條件下進(jìn)行,將水加進(jìn)雞糞稀釋 TS 濃度,這也是工程普遍采用的方式,可實(shí)現(xiàn)反應(yīng)器 的快速啟動(dòng)以及后期的穩(wěn)定運(yùn)行。由于雞糞有著較 高的固體含量,更適合于高固體厭氧消化技術(shù),但增 加雞糞進(jìn)料 TS 濃度勢(shì)必增加反應(yīng)器發(fā)酵液氨氮濃 度 ,實(shí) 現(xiàn) 高 固 體 雞 糞 厭 氧 處 理 仍 然 存 在 較 大 的 挑 戰(zhàn)[32] 。Bi 等[33, 34] 研究發(fā)現(xiàn),在雞糞固體濃度為 15%, 水 力 停 留 時(shí) 間(HRT)為 20~30 d,甲 烷 產(chǎn) 率 僅 為 0. 16~0. 29 L/g,并且有較高的有機(jī)酸殘余。雞糞中 溫厭氧消化系統(tǒng)的甲烷轉(zhuǎn)化率會(huì)隨著有機(jī)負(fù)荷和總 氨氮(TAN)的升高而受到抑制,在有機(jī)負(fù)荷達(dá)到 8. 5 kg TS(/ m3 ·d)以上,或 TAN 濃度達(dá)到 10000 mg/L 以上 時(shí),系統(tǒng)幾乎不再產(chǎn)氣直至崩潰[35] 。
2 環(huán)境因子對(duì)厭氧消化產(chǎn)甲烷體系的影響
2. 1 溫度對(duì)微生物代謝活動(dòng)的影響
溫度是影響厭氧消化效率的關(guān)鍵參數(shù),對(duì)微生 物的生長(zhǎng)速度和代謝有著重要影響,進(jìn)而關(guān)系到厭 氧消化體系中產(chǎn)甲烷性能。根據(jù)厭氧微生物適合生 存 的 環(huán) 境 溫 度 不 同 ,可 以 在 低 溫(15~25 ℃)、中 溫 (35~40 ℃)和高溫(50~60 ℃)條件下進(jìn)行。根據(jù)表 2 總結(jié)的產(chǎn)甲烷古菌生長(zhǎng)環(huán)境參數(shù)可知:多數(shù)產(chǎn)甲烷 古菌的最佳生長(zhǎng)溫度為 30~40 ℃的中溫環(huán)境,低溫和 高溫對(duì)大多數(shù)厭氧微生物而言為極端環(huán)境,不適宜 生長(zhǎng)[36, 37] 。
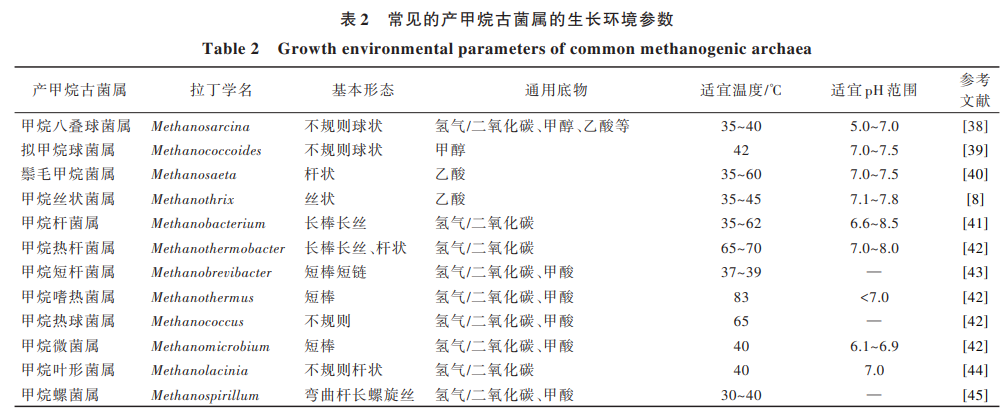
相比中溫環(huán)境,當(dāng)溫度降低至 10 ℃時(shí),系統(tǒng)內(nèi)氫 型產(chǎn)甲烷古菌甲烷桿菌目(Methanobacteriales)、乙酸 型產(chǎn)甲烷古菌甲烷八疊球菌目(Methanosarcinales)菌 群豐度均降低,系統(tǒng)內(nèi) VFAs 升高,甲烷產(chǎn)率有所下 降[46] 。王華等[47] 對(duì)比不同溫度條件下厭氧發(fā)酵的試 驗(yàn)結(jié)果發(fā)現(xiàn),在 10~30 ℃范圍內(nèi),產(chǎn)甲烷古菌的產(chǎn)氣 能力隨著溫度的降低而減小,并且其日產(chǎn)氣量的最 高峰也隨之往后推遲。當(dāng)溫度超過(guò) 40 ℃時(shí),微生物 菌群中的物種數(shù)量會(huì)急劇下降,47 ℃可能是中溫厭 氧產(chǎn)甲烷古菌生長(zhǎng)的上限[48] 。
在中溫厭氧消化過(guò)程中,甲烷的生成可經(jīng)由乙 酸營(yíng)養(yǎng)和氫營(yíng)養(yǎng)型產(chǎn)甲烷途徑實(shí)現(xiàn),但高溫厭氧消 化過(guò)程則主要通過(guò)氫營(yíng)養(yǎng)型產(chǎn)甲烷途徑產(chǎn)甲烷[49]。 雖然高溫環(huán)境下的厭氧微生物群落多樣性較低,但 比中溫卻有更高的有機(jī)物降解效率,在短時(shí)間內(nèi)對(duì) 寄生蟲(chóng)卵的殺滅率較高,有更高的產(chǎn)氣量。然而,高 溫厭氧消化能耗大、能源凈輸出降低、運(yùn)行成本較 高。因此,考慮到產(chǎn)甲烷的效率與反應(yīng)器運(yùn)行能耗 之間的平衡,且多樣性水平高的微生物菌群具有更 高的抗壓性和彈性,通常建議畜禽糞污厭氧消化產(chǎn) 生甲烷的溫度為 35~37 ℃[50] 。
2. 2 pH 對(duì)產(chǎn)酸代謝途徑的影響
pH 的動(dòng)態(tài)變化會(huì)引起微生物群落結(jié)構(gòu)的變化, 對(duì)系統(tǒng)穩(wěn)定性起著至關(guān)重要的作用。產(chǎn)甲烷古菌對(duì) pH 值極為敏感,當(dāng)環(huán)境 pH 值維持在 6. 5~7. 8 時(shí),對(duì) 大多數(shù)產(chǎn)甲烷古菌生存比較適宜。根據(jù)圖 2 厭氧消 化產(chǎn)甲烷代謝過(guò)程可知:約 70% 甲烷是產(chǎn)甲烷古菌 以乙酸為底物代謝的結(jié)果,目前為止,乙酸營(yíng)養(yǎng)型產(chǎn) 甲烷古菌只發(fā)現(xiàn)了甲烷八疊球菌屬 Methanosarcina 和 甲烷絲狀菌屬 Methanothrix,它們的最適 pH 范圍為 6. 8~7. 2。而水解與產(chǎn)酸菌對(duì) pH 值有較大范圍的適 應(yīng)性,大多數(shù)產(chǎn)酸菌群在 pH 值為 5. 0~8. 5 內(nèi)生長(zhǎng)良 好[51]。當(dāng)厭氧消化體系中 pH 值為 5. 5 時(shí),產(chǎn)甲烷速 率下降 80%;pH 值<5. 0 時(shí),厭氧體系幾乎不再產(chǎn)生 甲 烷 。 Sun 等[52]研 究 發(fā) 現(xiàn) ,當(dāng) pH 值 為 5. 1 時(shí) ,幾 乎 90%的乙酸營(yíng)養(yǎng)型甲烷菌和氫營(yíng)養(yǎng)型甲烷菌在 6 d 的培養(yǎng)過(guò)程中從培養(yǎng)物中消失,低 pH 環(huán)境提高了產(chǎn) 甲烷微生物的死亡速度,且這種抑制是不可逆的。 因此,pH 也是判斷厭氧消化產(chǎn)甲烷反應(yīng)是否正常進(jìn) 行的重要指標(biāo)。
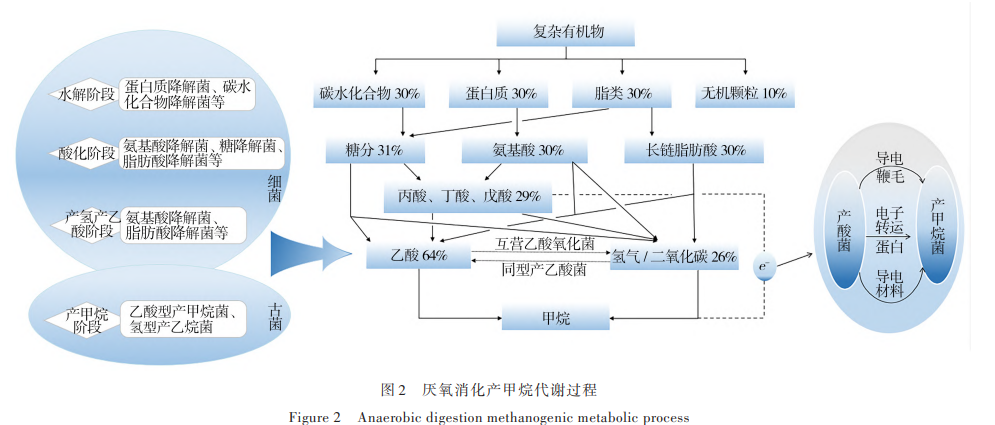
pH 可以改變厭氧消化過(guò)程中產(chǎn)酸代謝途徑,導(dǎo) 致 VFAs 的種類(lèi)和含量產(chǎn)生很大變化[53]。產(chǎn)酸代謝 過(guò)程中所產(chǎn)生的乙酸鹽、丙酸酯與丁酸酯的比例通 常是 75∶15∶10 至 40∶40∶20 [54]。Jiang 等[55]研究發(fā)現(xiàn) 當(dāng) pH 值為 5. 0 時(shí),產(chǎn)酸消化的主要產(chǎn)物是乙酸;但當(dāng) pH 值為 6. 0 或 7. 0 時(shí),主要產(chǎn)物則是丁酸。當(dāng)厭氧發(fā) 酵體系具有良好的緩沖能力時(shí),局部的 VFAs 濃度變 化過(guò)高不會(huì)顯著降低體系的 pH,對(duì)甲烷轉(zhuǎn)化效率不 會(huì)有顯著影響,系統(tǒng)中產(chǎn)氫產(chǎn)乙酸菌群和產(chǎn)甲烷古 菌群代謝動(dòng)態(tài)平衡在此起著關(guān)鍵作用。
2. 3 氨氮對(duì)厭氧消化系統(tǒng)的影響
在厭氧消化過(guò)程中,氨氮是微生物生長(zhǎng)所必需 的營(yíng)養(yǎng)物質(zhì),主要由發(fā)酵原料中的含氮有機(jī)化合物 降解而產(chǎn)生,如蛋白質(zhì)、氨基酸、尿素、核酸等,并以 NH+ 4 和 NH3 (FAN)分 子 的 形 態(tài) 存 在 ,二 者 之 和 為 TAN[56] 。氨氮在一定程度上可中和厭氧消化過(guò)程中所產(chǎn)生的有機(jī)酸,保證厭氧消化系統(tǒng)具有足夠的緩 沖能力,維持了細(xì)胞生長(zhǎng)所必需的中性 pH 條件,從而 提高厭氧消化系統(tǒng)的穩(wěn)定性。但糞污在厭氧消化過(guò) 程中,具有含氮高的特點(diǎn),過(guò)量的氨氮會(huì)對(duì)厭氧消化 產(chǎn)生抑制,影響厭氧消化系統(tǒng)的性能,甚至導(dǎo)致其失 穩(wěn)[57] 。厭氧微生物對(duì)原料的 C/N 也有一定的要求,碳 的消化速率比氮的速率快 30~35 倍,C/N 的最佳范圍 在理論上在 30~35 之間,通過(guò)加碳或氮來(lái)改變體系中 的碳氮比,以維持厭氧消化反應(yīng)的良好運(yùn)轉(zhuǎn)。
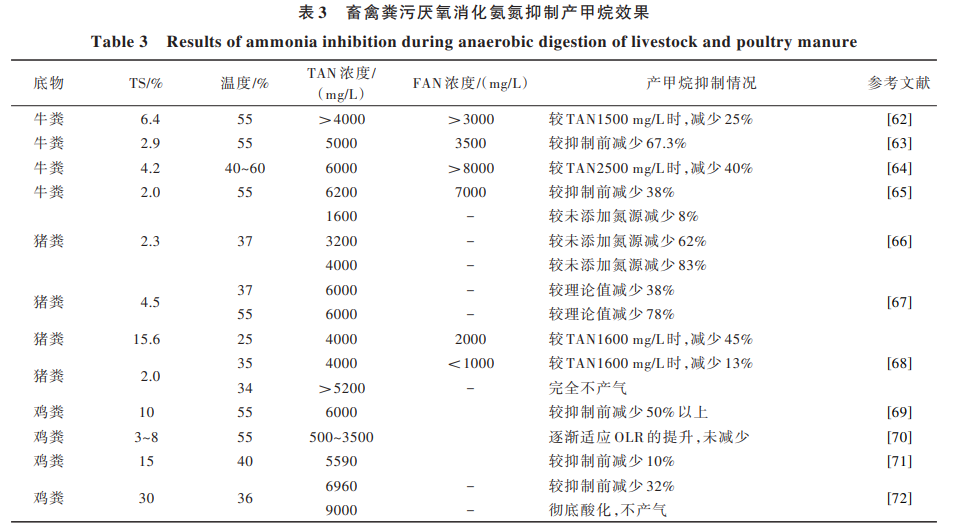
乙 酸 營(yíng) 養(yǎng) 型 產(chǎn) 甲 烷 古 菌 Methanosaeta 和 Methanosarcina 對(duì)氨氮更敏感,而嗜氫產(chǎn)甲烷古菌具 有較好的氨氮耐受性。當(dāng)氨氮濃度為 1300 mg/L 時(shí), 乙酸營(yíng)養(yǎng)型產(chǎn)甲烷古菌 Methanosarcina 開(kāi)始受到抑 制[58] 。當(dāng)氨氮濃度升高至 3500 mg/L 以上時(shí),嗜氫產(chǎn) 甲烷古菌 Methanoculleus 也會(huì)受到不同程度的氨抑 制[59, 60]。孟曉山等[61]研究發(fā)現(xiàn),豬糞厭氧消化系統(tǒng) 中氨氮超過(guò) 4000 mg/L,將促使產(chǎn)甲烷古菌群結(jié)構(gòu)發(fā) 生顯著變化,乙酸營(yíng)養(yǎng)型產(chǎn)甲烷古菌 Methanosaeta 逐 漸被 Methanosarcina 代替,而氫利用型產(chǎn)甲烷古菌屬 中 Methanospirillum 的優(yōu)勢(shì)性逐漸被 Methanoculleus 和 Methanomassiliicoccus 取代,高濃度氨氮也會(huì)促使產(chǎn)甲 烷途徑由乙酸利用型為主向氫利用型為主轉(zhuǎn)變。在 畜禽糞污厭氧消化過(guò)程中,氨氮抑制產(chǎn)甲烷效果的 相關(guān)情況總結(jié)見(jiàn)表 3。
3 進(jìn)料負(fù)荷和進(jìn)料方式對(duì)厭氧消化產(chǎn)甲烷效能的 影響
3. 1 水力停留時(shí)間對(duì)厭氧消化產(chǎn)甲烷效能的影響
有機(jī)負(fù)荷率(OLR)是運(yùn)行厭氧消化系統(tǒng)的重要 工程控制參數(shù),通過(guò)調(diào)整 HRT 可改變系統(tǒng)的有機(jī)負(fù) 荷,從而平衡畜禽糞污厭氧消化產(chǎn)酸速率與產(chǎn)甲烷 效率,實(shí)現(xiàn)最佳的產(chǎn)氣性能[73]。厭氧消化反應(yīng)器產(chǎn) 氣量會(huì)隨進(jìn)料有機(jī)負(fù)荷的增加而加大,但當(dāng) HRT 縮 短到一定程度,則會(huì)因負(fù)荷過(guò)高而造成有機(jī)揮發(fā)酸 的積累,尤其是丙酸,對(duì)厭氧消化過(guò)程產(chǎn)生抑制,從 而導(dǎo)致產(chǎn)氣量降低[74] 。HRT 從 10 d 減少到 7. 5 d,促 進(jìn)了甲烷八疊球菌屬 Methanosarcina 的生長(zhǎng),相對(duì)甲 烷絲狀菌屬 Methanothrix 具有優(yōu)勢(shì)[75]。單純的糞污 厭 氧 消 化 效 率 相 對(duì) 較 低 ,抗 降 解 成 分 需 要 較 高 的 HRT。通過(guò)與餐廚垃圾、粗甘油、秸稈等物料共消化 可有效提高甲烷產(chǎn)量。在牛糞與餐廚垃圾共消化的 研究中,發(fā)現(xiàn)縮短 HRT(25,20,15,10,7,5,4 d)會(huì)減 少甲烷轉(zhuǎn)化率,系統(tǒng)中擬桿菌門(mén)(Bacteroidetes)和厚 壁菌門(mén)(Firmicutes)成為優(yōu)勢(shì)菌,醋酸合養(yǎng)氧化菌(主 要是 Pelotomaculum 和 Pseudothermotoga)的比例明顯 增加;優(yōu)勢(shì)產(chǎn)甲烷古菌由乙酸營(yíng)養(yǎng)型產(chǎn)甲烷古菌轉(zhuǎn) 變?yōu)槭葰湫彤a(chǎn)甲烷古菌[76, 77]。在不同的 HRT 下,不 同類(lèi)型底物的厭氧消化系統(tǒng)中產(chǎn)甲烷古菌變化情況 見(jiàn)表 4。

為提高畜禽糞污的處置效率,常見(jiàn)的減少 HRT 的方法有混合攪拌和預(yù)處理。混合攪拌可使消化物料混合和分布均勻,防止物料在發(fā)酵罐底部沉淀,增 大反應(yīng)物與微生物的接觸面積,有利于保證良好的 消化速度,從而提高消化產(chǎn)氣量。Karim 等[80] 研究發(fā) 現(xiàn),機(jī)械攪拌、沼氣回流攪拌、出水回流攪拌和未攪 拌相比,厭氧消化的產(chǎn)氣量分別提高了 22%、15% 和 29%。預(yù)處理方法中熱水解法應(yīng)用最為廣泛,糞污預(yù) 處理后的營(yíng)養(yǎng)成分更容易被微生物利用,厭氧消化 能力提高,停留時(shí)間也相應(yīng)減少[81]。宋曉聰?shù)龋?2]將 牛糞于 70 ℃處理 3 d 后進(jìn)行厭氧消化,發(fā)現(xiàn)其產(chǎn)氣量 較未處理的牛糞提高了 210%,有機(jī)組分降解和木質(zhì) 素含量降低是產(chǎn)氣性能提高的原因。
3. 2 不同進(jìn)料方式對(duì)厭氧消化產(chǎn)甲烷效能的影響
不同的進(jìn)料方式可能造成微生物群落差異,進(jìn) 而影響產(chǎn)氣率。在較高的 OLR 下,低進(jìn)料頻率可能 在短時(shí)間內(nèi)對(duì)系統(tǒng)造成較重的負(fù)荷沖擊,高進(jìn)料頻 次可以有效降低消化系統(tǒng)的負(fù)荷沖擊,使系統(tǒng)產(chǎn)生 的 VFAs 能夠被及時(shí)消耗[83]。序批式間歇流反應(yīng)器 (SBR)的低進(jìn)料頻次會(huì)導(dǎo)致底物梯度隨時(shí)間變化,初 始底物濃度較高。相反,連續(xù)流反應(yīng)器(CFR)的長(zhǎng)進(jìn) 料時(shí)間會(huì)促使系統(tǒng)時(shí)刻保持穩(wěn)定和低底物濃度狀 態(tài)[84] 。Kine 等[85] 研究發(fā)現(xiàn),每天進(jìn)料 1 次時(shí),丙酸的 濃度峰值為 2500 mg/L,而每 2. 4 h 進(jìn)料 1 次時(shí),丙酸 濃 度 始 終 保 持 在 40 mg/L 以 下 。 甲 烷 八 疊 球 菌 屬 Methanosarcina 在間歇式進(jìn)料的情況下表現(xiàn)出一定優(yōu) 勢(shì),而甲烷絲狀菌屬 Methanothrix 則表現(xiàn)出相反的行 為[86]。在較低 OLR 下,不同進(jìn)料頻次均未產(chǎn)生較重 的負(fù)荷沖擊,酸化現(xiàn)象一般不會(huì)出現(xiàn);且無(wú)論 HRT 或 進(jìn)料方式的如何變化,甲烷絲狀菌屬 Methanothrix 均 為主要的產(chǎn)甲烷古菌屬[87] 。
4 外部添加材料對(duì)厭氧消化產(chǎn)甲烷效能的提升
4. 1 投加功能菌劑對(duì)厭氧消化產(chǎn)甲烷效能的提升
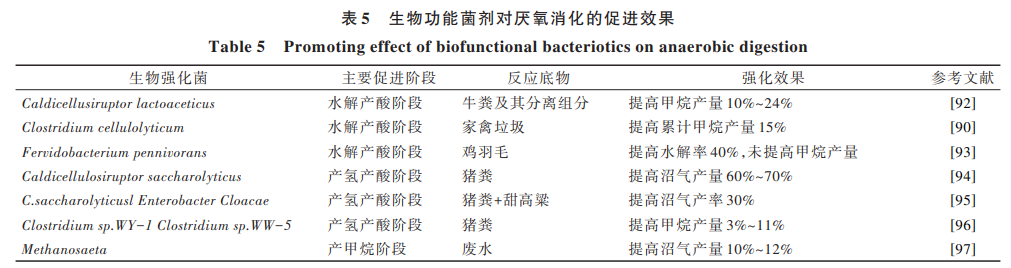
向反應(yīng)系統(tǒng)中投入具有一定功能的微生物功能 菌劑,能夠在很大程度上提高生化效率,達(dá)到加快厭 氧系統(tǒng)啟動(dòng)時(shí)間、縮短 HRT、增加難降解原料利用 率、促進(jìn)酸敗系統(tǒng)的恢復(fù)、應(yīng)對(duì)氨抑制等目標(biāo)[88] 。根 據(jù)目標(biāo)產(chǎn)物的不同,可將其劃分為水解產(chǎn)酸、產(chǎn)氫產(chǎn) 乙酸、產(chǎn)甲烷階段的生物強(qiáng)化,其促進(jìn)效果見(jiàn)表 5。 由于畜禽糞污中含有大量的木質(zhì)纖維素,有效發(fā)酵 成分不易降解,對(duì)水解產(chǎn)酸階段生物強(qiáng)化,促進(jìn)木質(zhì) 纖維原料的水解,為厭氧消化可提供更多易降解的 底物。Henrik 等[89] 研究發(fā)現(xiàn),向牛糞及其分離組中投 加乳酸桿菌 Caldicellulosiruptor lactoaceticus 可以使得 甲 烷 產(chǎn) 率 提 高 10%~24%。 然 而 ,Costa 等[90]研 究 發(fā) 現(xiàn),盡管在高溫厭氧發(fā)酵畜禽糞便體系中投加熱線 梭 菌 Clostridium thermocellum 及 解 糖 纖 維 素 分 解 菌 Caldicellulosiruptor saccharolyticus,雖然原料水解率提 高了,但甲烷的產(chǎn)量并沒(méi)有明顯增多,這是由于產(chǎn)甲 烷速率跟不上纖維素水解產(chǎn)酸菌速率,導(dǎo)致 VFAs 累 積,抑制了產(chǎn)甲烷。通過(guò)對(duì)水解產(chǎn)酸階段的生物強(qiáng) 化,向系統(tǒng)中投加產(chǎn)氫菌,所產(chǎn)的沼氣中氫氣含量并 未增加,即甲烷與氫氣組分比例無(wú)明顯減小,說(shuō)明投 加產(chǎn)氫菌所增加的氫氣是以中間產(chǎn)物存在的。氫氣 作為產(chǎn)甲烷古菌的底物,被進(jìn)一步用于產(chǎn)甲烷。此 外,產(chǎn)氫菌的生物強(qiáng)化還可使甲烷菌的數(shù)量增加,即 氫營(yíng)養(yǎng)型產(chǎn)甲烷古菌在數(shù)量上得到了強(qiáng)化[91] 。
根據(jù)圖 2 厭氧消化產(chǎn)甲烷代謝過(guò)程可知:產(chǎn)酸細(xì) 菌和產(chǎn)甲烷古菌之間存在種間電子轉(zhuǎn)移(IET),該方 式是菌屬之間互養(yǎng)代謝產(chǎn)甲烷極其重要的環(huán)節(jié)。由于較快的水解發(fā)酵產(chǎn)酸和較慢的產(chǎn)甲烷之間的不平 衡,從而導(dǎo)致畜禽糞污厭氧消化處理負(fù)荷只能維持 在相對(duì)較低效能,產(chǎn)甲烷效率受到了限制[98] 。IET 產(chǎn) 甲烷途徑是微生物菌群降低熱力學(xué)能壘、實(shí)現(xiàn)互養(yǎng) 代謝快速產(chǎn)甲烷的核心過(guò)程。互養(yǎng)代謝產(chǎn)甲烷過(guò)程 除了常見(jiàn)已知的種間氫轉(zhuǎn)移[99] 和種間甲酸轉(zhuǎn)移[100] 之 外,美國(guó)馬薩諸塞大學(xué) Derek Lovley 教授團(tuán)隊(duì)[101]于 2010 年在國(guó)際上首次報(bào)道了電活性微生物硫還原地 桿 菌 Geobacter sulfurreducens 和 金 屬 還 原 地 桿 菌 Geobacter metallireducens 之間存在直接種間電子傳遞 (DIET)過(guò)程,這種代謝方式不需要電子穿梭體,可以突 破傳遞過(guò)程的速率限制,能夠以更高的電子轉(zhuǎn)移效率 或較低的能量消耗直接將電子轉(zhuǎn)移至二氧化碳,將二 氧化碳還原為甲烷,可極大提升產(chǎn)甲烷速率。目前已 發(fā)現(xiàn) 2 類(lèi)種間直接電子傳遞形式,一類(lèi)是依靠細(xì)胞外 膜上的c型細(xì)胞色素,另一類(lèi)是通過(guò)導(dǎo)電菌毛(pili)[102] 。
通過(guò)對(duì)參與 DIET 產(chǎn)甲烷途徑的互養(yǎng)代謝微生物 群落結(jié)構(gòu)進(jìn)行厭氧消化調(diào)控,增加可參與 DIET 過(guò)程 的微生物豐度,進(jìn)而提高產(chǎn)甲烷效率。Jiang 等[92]研 究發(fā)現(xiàn)通過(guò)添加富集的丙酸降解產(chǎn)甲烷培養(yǎng)基,培 養(yǎng)基中所含優(yōu)勢(shì)菌屬為甲烷絲狀菌屬 Methanothrix (66. 15%~94. 70%)和 甲 烷 桿 菌 屬 Methanobacterium (4. 81%~33. 56%),相比與非生物強(qiáng)化組,系統(tǒng)中甲 烷絲狀菌屬 Methanothrix 的豐度超過(guò) 80%,有機(jī)負(fù)荷 和沼氣產(chǎn)量分別提高 8 倍和 12 倍。目前報(bào)道的確認(rèn) 可以參與直接種間電子傳遞的菌屬(表 6)可知:供電 子微生物多為具有胞外電子傳遞能力的微生物,如 地桿菌屬 Geobacter,而受電子的產(chǎn)甲烷古菌則主要是 可以利用乙酸鹽的鬃毛甲烷菌屬 Methanosaeta 和甲 烷八疊球菌屬 Methanosarcina。

微生物功能菌劑投加成功的關(guān)鍵是能否在系統(tǒng) 中持久穩(wěn)定地發(fā)揮作用,所投加的微生物功能菌繁 殖速度需高于其被土著菌吞噬及流失的速度,保證 生物強(qiáng)化菌劑在被強(qiáng)化體系內(nèi)的生物凈持有量。為 增加生物強(qiáng)化菌劑在被強(qiáng)化體系中的穩(wěn)定性,可將 生物強(qiáng)化菌劑固定化在載體上再進(jìn)行投加,固定化 的載體主要有沸石、海泡石等[109, 110] 。對(duì)于連續(xù)流系 統(tǒng),在不斷地進(jìn)出料沖擊及土著微生物的競(jìng)爭(zhēng)下,微 生物功能菌劑容易逐漸被淘汰,定期多次投加生物 強(qiáng)化菌劑效果優(yōu)于一次性投加[111] 。
4. 2 投加導(dǎo)電材料對(duì)厭氧消化產(chǎn)甲烷效能的提升
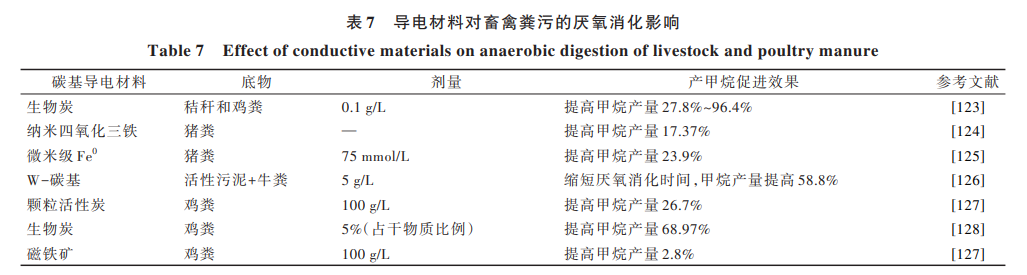
采用導(dǎo)電材料強(qiáng)化 DIET 產(chǎn)甲烷途徑的畜禽糞污厭氧處理技術(shù),是提高畜禽糞污資源利用的有效途 徑和研究熱點(diǎn)。在厭氧系統(tǒng)中投加外源導(dǎo)電材料 (如顆粒活性炭[112]、碳布[113]、生物炭[114]、石墨烯[115]、 磁鐵礦[116] 、磁性顆粒活性炭[117] 、聚苯胺[118] 等)可強(qiáng)化 甲烷八疊球菌屬 Methanosarcina 等產(chǎn)甲烷功能菌參與 DIET 過(guò)程,進(jìn)而提高厭氧體系甲烷轉(zhuǎn)化效率,其具體 影響效果見(jiàn)表 7。外源導(dǎo)電材料具有高導(dǎo)電性、化學(xué) 穩(wěn)定性、吸附能力強(qiáng)等特點(diǎn),能夠作為生物導(dǎo)電連接 重要補(bǔ)充;同時(shí),不需要微生物消耗能量以合成生物 電連接的自身導(dǎo)電結(jié)構(gòu),微生物可以利用更多能量 于代謝作用。Pan 等[119] 研究發(fā)現(xiàn),添加生物炭可以提 高雞糞、牛糞及其混合物的產(chǎn)氣潛力,并能提高所 產(chǎn)沼氣的甲烷含量,通過(guò)吸附作用,降低氨氮和硫 化氫的含量。Lee 等[120, 121]研究發(fā)現(xiàn)在系統(tǒng)中投加 生 物 炭 和 活 性 炭 后 ,其 表 面 富 集 了 鬃 毛 甲 烷 菌 屬 Methanosaeta、甲 烷 螺 菌 屬 Methanospirillum 和 甲 烷 桿 菌 屬 Methanobacterium 等 產(chǎn) 甲 烷 古 菌 。 Zeng 等[122]探究了碳基導(dǎo)電材料負(fù)載核黃素后對(duì)畜禽糞 污 厭 氧 消 化 的 影 響 ,通 過(guò) 投 加 核 黃 素 改 變 了 微 生 物 群 落 結(jié) 構(gòu) 并 提 高 了 與 DIET 相 關(guān) 的 功 能 基 因 豐 度 ,最 終 提 高 了 反 應(yīng) 器 的 容 積 產(chǎn) 氣 率 和 有 機(jī) 物 去 除率。
4. 3 投 加 非 導(dǎo) 電 材 料 對(duì) 厭 氧 消 化 產(chǎn) 甲 烷 效 能 的 提升
在厭氧系統(tǒng)中投加外源非導(dǎo)電材料(如尼龍纖 維[129] 、聚乙烯[130] 、堿性樹(shù)脂[131] 等)也可強(qiáng)化厭氧體系 產(chǎn)甲烷效率。非導(dǎo)電材料能夠提升甲烷產(chǎn)率主要是 因其比表面積大,易于吸附定植相關(guān)功能微生物,如 厭氧消化細(xì)菌、產(chǎn)甲烷古菌等,使其能夠高效地參與 到厭氧消化過(guò)程,部分載體還具有吸附能力、離子交 換能力、優(yōu)化菌群結(jié)構(gòu)等特點(diǎn)[132] 。不同材質(zhì)類(lèi)型、粒 徑及濃度對(duì)厭氧消化的影響存在較大的差異,還有 可能存在抑制厭氧消化的情況。Zhang 等[133] 發(fā)現(xiàn)在 55,65 ℃下,聚乙烯塑料的存在能使奶牛糞污厭氧消 化的甲烷產(chǎn)量分別提高 8. 4%、41. 2%,COD 降解率分 別提高到 52. 8% 和 52. 4%。而厭氧消化系統(tǒng)中塑料 含量超過(guò) 0. 2 g/L 對(duì)厭氧消化有不良影響,甲烷產(chǎn)量 減少了 17. 9%~19. 3%,這是由于厭氧消化系統(tǒng)中存 在的微塑料物質(zhì)影響了微生物種群的豐度和結(jié)構(gòu), 使得甲烷桿菌屬 Methanobacterium 和鬃毛甲烷菌屬 Methanosaeta 的豐度降低[134] 。
5 結(jié)束語(yǔ)
厭氧消化技術(shù)是處置畜禽糞污有效手段,通過(guò) 對(duì)比分析牛糞、豬糞、雞糞的糞污特性,牛糞因含有 較多不易降解有機(jī)組分進(jìn)而產(chǎn)甲烷潛力最低,雞糞 因含有較多營(yíng)養(yǎng)物質(zhì)進(jìn)而產(chǎn)甲烷潛力最高。適宜的 溫度、pH、氨氮等環(huán)境條件是保障畜禽糞污厭氧消化 穩(wěn)定性運(yùn)行的基礎(chǔ)。基于系統(tǒng)產(chǎn)甲烷微生物的代謝 環(huán)境要求,pH、氨氮變化也成為判斷厭氧消化產(chǎn)甲烷 反應(yīng)是否正常進(jìn)行的重要指標(biāo)。不同的 HRT、進(jìn)料 方式能夠改變系統(tǒng)產(chǎn)甲烷古菌類(lèi)型,也會(huì)對(duì)禽糞污 厭氧消化產(chǎn)甲烷效能產(chǎn)生影響。基于厭氧消化影響 的關(guān)鍵因素探究,通過(guò)功能菌劑、導(dǎo)電材料等材料的 投加,調(diào)控系統(tǒng)功能性微生物群落豐度,強(qiáng)化電活性 微生物之間的直接種間電子傳遞,促進(jìn)厭氧消化高 負(fù)荷運(yùn)行,進(jìn)而提升產(chǎn)甲烷效能。未來(lái)的畜禽糞污 厭 氧 消 化 產(chǎn) 甲 烷 效 能 提 升 研 究 ,應(yīng) 重 點(diǎn) 關(guān) 注 以 下 方面:
1)產(chǎn)甲烷古菌的 DIET 代謝路徑機(jī)制研究。從細(xì) 胞水平對(duì)產(chǎn)甲烷古菌的 DIET 代謝路徑進(jìn)行優(yōu)化提 速,是提高系統(tǒng)有機(jī)質(zhì)轉(zhuǎn)化率以及容積負(fù)荷的重點(diǎn) 突破口。產(chǎn)甲烷速率低于產(chǎn)酸速率時(shí),有機(jī)酸在體 系中積累造成系統(tǒng)酸化,酸積累導(dǎo)致厭氧發(fā)酵系統(tǒng) 酸化的抑制作用。隨著微生物菌群種間電子傳遞研 究的深入,產(chǎn)甲烷功能菌株、導(dǎo)電材料也逐漸成為厭 氧發(fā)酵失穩(wěn)調(diào)控策略的研究熱點(diǎn),如何構(gòu)建特異性產(chǎn)甲烷功能菌株、改性導(dǎo)電材料來(lái)提高厭氧發(fā)酵系 統(tǒng) 穩(wěn) 定 性 和 處 理 效 率 ,成 為 理 論 研 究 亟 待 解 決 的 問(wèn)題。
2)智能化調(diào)控的厭氧消化裝備研發(fā)。為精確控 制厭氧消化過(guò)程中的關(guān)鍵參數(shù),如溫度、pH 值、攪拌 強(qiáng)度、進(jìn)料負(fù)荷等,創(chuàng)造最適宜微生物生長(zhǎng)和產(chǎn)甲烷 的環(huán)境條件,可通過(guò)先進(jìn)的傳感器技術(shù)和在線監(jiān)測(cè) 設(shè)備,實(shí)時(shí)監(jiān)測(cè)厭氧消化過(guò)程中的各項(xiàng)參數(shù),建立智 能化的控制系統(tǒng),進(jìn)而實(shí)現(xiàn)對(duì)消化過(guò)程的精準(zhǔn)調(diào)控。 基于自動(dòng)化運(yùn)行控制,及時(shí)調(diào)整工藝參數(shù),可避免因 參數(shù)波動(dòng)導(dǎo)致的系統(tǒng)不穩(wěn)定和產(chǎn)甲烷效率下降。

相關(guān)問(wèn)題
環(huán)境相關(guān)核刊
環(huán)境類(lèi)相關(guān)問(wèn)題解答
- 環(huán)境科學(xué)方向中科院SCI一二區(qū)TOP刊匯總
- 錄用率高達(dá)75%,環(huán)境類(lèi)sci中科院一二區(qū)期刊
- 已投修改后錄用的環(huán)境科學(xué)與生態(tài)學(xué)sci期刊
- 北大核心+CSCD+AMI擴(kuò)展 《環(huán)境科學(xué)研究》投稿格式及要求
- 2025年上海市工程系列生態(tài)環(huán)境保護(hù)專(zhuān)業(yè)評(píng)中級(jí)職稱(chēng)論文的要求
- 雙核心《安全與環(huán)境工程》《環(huán)境工程技術(shù)學(xué)報(bào)》論文投稿模板(可下載)
- IF:11+,審稿快,環(huán)境科學(xué)與生態(tài)學(xué)1區(qū)top期刊,無(wú)版面費(fèi)
- 幾乎不退稿的SCI期刊:水利、土木、環(huán)境
- 環(huán)境科學(xué)與生態(tài)方向SCI二(Q2)區(qū)期刊,IF:2.4
- 環(huán)境科學(xué)與生態(tài)學(xué)領(lǐng)域sci期刊列表(348本)